
Natural selection in mycorrhizal symbioses
By: Dr. César Marín, Full Professor, Universidad Santo Tomás, Chile | Editor-in-Chief, International Mycorrhiza Society Newsletter
Many biologists hold a strong gene-centric view of evolution: they define evolution as a change in the frequency of alleles. However, many widespread processes, particularly in plants and their microbiomes, complicate this definition: epistasis – where the effect of a gene on the phenotype depends on a complex network of interactions; epigenetic inheritance (heritable changes that do not alter DNA sequences), and horizontal gene transfer between hosts microbes complicate a gene-centric view of evolution, where there is a simple correspondence between genotype and phenotype (one or few non-interacting genes explain one trait). In addition, conceptual and empirical developments over the last century, show that natural selection occurs across multiple levels of the biological hierarchy. In particular, Multilevel Selection (MLS) theory argues that natural selection operates simultaneously in multiple levels of the biological hierarchy (from molecules to communities). This is because at the individual level, selfish individuals outcompete cooperative individuals, but cooperative groups outcompete selfish groups. For example, Darwin argued that within a tribe, individuals who behave more selfishly will have advantages and out-compete the more cooperative individuals, but tribes with individuals “always ready to give aid to each other and sacrifice themselves for the common good, would be victorious over most other tribes; and this would be natural selection” (Darwin, 1871; The Descent of Man, p. 166). Other common examples of MLS theory include the evolution of eusocial insects such as bees and ants -it would not make sense to think that natural selection operates solely at each individual insect level, cancer (where natural selection operates at the organismal and cellular levels), and holobiont (an animal or plant host and its fitness-aligned microbiome) evolution. MLS has long been used in poultry science, where artificially selecting entire cages of less hostile hens produces more eggs than selecting the individual hens that produce more eggs. Please find here a simple explanation of this theory. During the 12th International Conference on Mycorrhiza, which took place in Manchester, UK, in August of 2024, Nancy Collins Johnson and I chaired a workshop dedicated to understand how MLS theory can be used to understand the mycorrhizal symbiosis, across multiple scales.
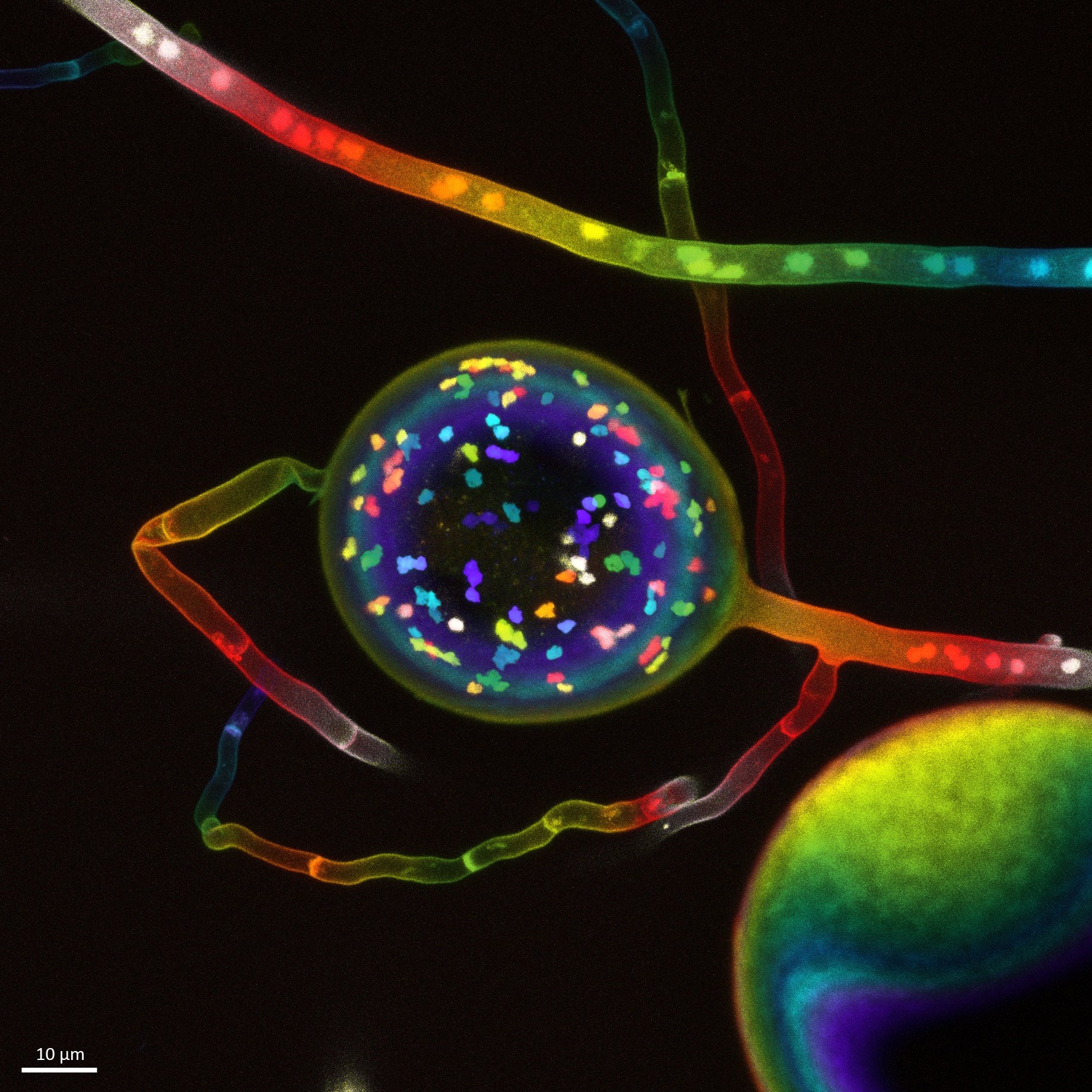
Fig. 1. Spore of an arbuscular mycorrhizal fungus containing multiple nuclei within (represented in different colors). Photo by: Vasilis Kokkoris, Franck Stefani, and Nicolas Corradi.
Hundreds of studies have recognized MLS in natural and laboratory populations, and it may be a useful model, particularly for understanding the evolution of arbuscular mycorrhizal (AM) symbioses. Historically, there has been a conflation of the units involved in natural selection (“units of selection” debate) and uncoupling them may be enlightening. We argue that recognizing three distinct units of natural selection may facilitate a MLS paradigm for the evolution of AM symbioses. One unit interacts with the environment and leads to differential fitness (interactors), the second unit generates inheritance (replicators/reproducers/reconstitutors), and the third unit manifests accumulated or optimized adaptations (manifestors of adaptations). A detailed explanation of these units of selection can be found here. These three different evolutionary functions do not need to occur at the same level of the biological hierarchy. For example, genes would typically constitute replicators, holobionts can be considered as reconstitutors (as their microbiome reconstitutes each generation), while individual organisms (but also groups of organisms, and holobionts and communities) can constitute an interactor. Typically, individual organisms are classical examples of manifestors of accumulated adaptations – as Darwin finches’ beaks, where the change is gradual and accumulates over time. Accumulated adaptations contrast with “product-of-selection” adaptations, like the industrial melanism in moths, where due to the industrial Revolution in the XIX century, it was noted that dark moths became more common. In such case, is the frequency of dark/white moths what changes, but not their biology. We suggest that an understanding of the types of adaptations through this tripartite framework of units of selection could help explain phenomena such as the unusual nuclear dynamics of AM fungi (as a single AM fungal spore can have hundreds of nuclei; Fig. 1), and the presence of a core, P-mineralizing microbiome in the AM fungal hyphosphere (ie. in the microbiome inhabiting the hyphae). This workshop explored how natural selection operates across the biological hierarchy in AM systems, from nuclei to groups of plant holobionts and ecosystems. It included five talks by: César Marín (Chile; introduction to this framework); Sander van Otterdijk (The Netherlands; selection at the nuclei level); Letian Wang (China; on a core functional microbiome in the AM fungal hyphosphere); James D. Bever (US; on plant-soil-feedbacks and the relationship of this concept with MLS theory); and Nancy Collins Johnson (US; on “Functional Team Selection” as a framework to understand how plants assemble their belowground microbiome) (Fig. 2).
We hope that this workshop will illuminate debates regarding individuality and holobionts in AM systems and help advance the development of theories and experimental systems to elucidate mechanisms of mycorrhizal evolution. This knowledge is necessary before AM symbioses can be effectively managed in agriculture and ecosystem restoration.
YouTube link of the workshop: https://youtu.be/vKDwfqW6Ask?si=2mpvwh0NWEDBslom
Website: https://cesar-marin.com/

Fig. 2. From left to right: Letian Wang, Sander van Otterdijk, César Marín, Nancy Collins Johnson, James D. Bever. Photo by: Patricia Silva-Flores.